CHAPTER 2
Sections of this chapter
![]()
CHAPTER 2
Sections of this chapter
![]()
There is no clear dividing line in terms of its operational properties between the purely chemical system we have called "a protocell" and a living cell. Both metabolize, grow, mutate, reproduce, and die. Metabolism refers to the property by which a self-reproducing system absorbs nutrients and turns them into parts of itself and useful energy. The protocells may appear completely passive in that their functions are completely determined by chemical laws, while cells, such as an amoeba, seem to act purposefully to seek out nutrients and metabolize them. However, arguments can be made that the apparent purposefulness of the amoeba in capturing and digesting nutrients is completely determined by physicochemical laws and its own physicochemical structure. Plants, which are much more evolved than amoebas, seem as physicochemically determined and passive as the protocells. Therefore, there may be no true or meaningful dividing line between matter and life. Both are manifestations of a single evolutionary process. But life seems to make choices while matter does not. More will be said of this in Chapter 5. First we will consider the more deterministic patterns of evolution.
The Pattern of Evolution (Preview)Tracing the evolutionary process from the earliest beginnings currently perceivable to the present, we see the following pattern emerge. Chaotic energy organizes into elementary particles; these organize into systems of particles (nucleons); the nucleons organize into systems of nucleons (atoms); the single atoms organize into large but simply connected systems of atoms (galaxies, stars and planets); within the stars more complex atoms are built from simpler atoms; within the planets, and to a lesser extent outside the planets, atoms organize into small but complexly connected sys-tems of atoms (molecules); the molecules organize into increasingly complexsystems of molecules until these systems are self-replicating; then evolutionby natural selection begins and leads to symbiotic systems of self-replicating molecules (the protocells) which through autopoiesis become first bacteria; then the bacteria organize into nucleated cells. We shall later show how this pattern is continued with cells organizing into autopoietic systems of cells (i.e., multicellular animals - the metazoa) and how these systems become increasingly complex until they are capable of consciously directing their own evolution through higher-order autopoiesis. These latter systems (human beings) then organize into complex interdependent symbiotic, but not yet autopoietic, societies.
We note the following pattern:
- (1)Each system incorporates all the properties and information of its subsystems plus some new properties and information of its own; the latter are epiphenomena of the new higher-order autopoiesis.
- (2)Higher systems cannot exist without lower systems.
- (3)The basic systems are composed of a central organizing nucleus with simpler interdependent subsystems in the periphery, e.g., atoms, galaxies, solar systems, cells and their nuclei, humans and the brain, human societies and their central governments.
- (4)Each time a new level of organization (autopoiesis) evolves, there occurs a new dimension of complexity, to be called dimensional quadrature (e.g.: nucleons becoming atoms, atoms becoming molecules, molecules becoming bacteria, bacteria becoming nucleated cells, cells becoming metazoa, metazoa becoming human).
- (5)As we will show later, this is a process which may have no end.
- (6)A common denominator is ever-increasing complexity and total coherent information content, which enables each system to use all its subsystems and related systems to synthesize ever more complex supersystems with ever more coherent information.
The Cellular SynthesisThe developmental steps leading from the first protocell to the final cell may have been as great as the steps leading from atom to protocell. The major steps in cellular development involved (1) the evolution of RNA, protein, and DNA within the cell, (2) the specialization of proteins into enzymes, cytoplasm, and the related subsystems, (3) the formation of a protective semipermeable membrane, (4) the development of a central nucleus as bacteria evolved into cells, and (5) directed locomotion.
It is almost certain that through natural selection a symbiotic interdependency developed between the nucleic acid and protein of the protocell until neither one of these components could survive without the other - the beginning of autopoiesis. The point at which autopoiesis came into being is the closest approximation to the point at which life as we know itstarted, although this is clearly somewhat arbitrary. The protocells would eventually replace all the autocatalyzing molecules, possibly by metabolizing them, and then begin competing against one another. Those protocells best able to assimilate nutrients and reproduce would replace the less efficient protocells, i.e., those that were not autopoietic. The evolutionary pressure was on increasing the capability for (1) obtaining nutrients and (2) reproducing. Those protocells which first developed the capability for movement had a distinct advantage in obtaining nutrients over the protocells which were dependent on currents of water for moving them to places where there were nutrients. If these protocells ever became stagnated in nutrient-poor pools, they would die out. Eventually protocells with the capability of movement predominated.
Movement could at first have been due to vibrations sent up in the outer parts of the protocell by the absence of nutrients. Some of the proteins would have mutated into vibratory molecules which had their vibrations damped by nutrients so that the protocells would always move in the direction of maximum nutrient concentration. The cilia are proteins, which have this property and will behave in this way independently of the rest of the cell. This gives the appearance of purposive action to a completely automatic chemical process. However, the information within the cell also modulates the cilia.
"Evolutionary pressure" refers to the propensity of natural selection to favor some mutations over others because of the current environmental opportunities that exist for those mutations. It is a pressure that pulls from the future rather than pushes from the past. Once the evolutionary pressure through natural selection was pulling the protocell toward symbiotic development, mutations which favored one subsystem of the protocell at the expense of another would not survive. Movement favored the entire protocell. However, increased efficiency in metabolism of nutrients in only one part (say, the outer covering) of the protocell would lead to asymmetric and deleterious growth. Therefore, fairly early in the evolution of the protocell a subsystem had to be developed for distributing nutrients to all parts of the protocells in a balanced way. This might have occurred at first by having a circulating homogeneous mixture of nutrients so that the nutrients were equally divided among all parts of the protocell. Eventually it was to be done through special metabolic pathways by specialized molecules through the process called photosynthesis.
PhotosynthesisThe protocells probably required fairly complex molecules such as amino acids and nucleotides for nutrients. As the protocells evolved, these nutrients, which were probably contained mainly in the structures of the older autocatalyzing molecules, became increasingly scarce. The only readilyavailable molecules then were relatively simple ones such as HCN, NH3, CH4 and CO2. The latter had now become a major constituent of the atmosphere and was probably a metabolic product of some of the early protocells. We recall that the building of complex molecules from simple molecules requires an input of energy. As the earth evolved there was ever less energy for this purpose except from sunlight, where the energy may have been increasing. But the sunlight is very diffused over the entire surface of the earth and it could not readily provide, except in the upper atmosphere from solar flares, those bursts of energy which are necessary to pull the simple molecules up the evolutionary ladder. These bursts probably continued to come from lightning, volcanic action, cosmic rays, meteors, background radiation, and the like impinging on concentrated pools of molecules. These events may still occur in the suboceanic volcanic vents. However, there is a fairly simple physicochemical process called photosynthesis for concentrating solar energy. The photosynthetic process has no survival value by itself but only when it is incorporated as part of a self-replicating system. Only a highly evolved protocell or clearly living cell would be likely to have the complex subsystems to support and utilize photosynthesis. However, once photosynthesis began, there was a literal explosion of life.
At this time, the atmosphere and the oceans were loaded with CO2 and there was very little free oxygen. Some oxygen had probably been produced from early times by the disassociation of water in the upper atmosphere as follows: 3H2O + photon 3H+ O3. The H escapes to outer space. This is probably how Mars, with much less gravity than earth, became depleted of water. The ozone forms a protective cover against high-energy ultraviolet rays and breaks down into oxygen: 2O 3O. Once a series of mutations began to give the protocell the capability for synthesizing nutrients from simple molecules and sunlight, it had an enormous advantage over its less chemically sophisticated relatives. Its progeny would expand and fill the entire biosphere. All non-photosynthetic protocells would tend to become extinct. Almost all life on earth is descended from these early photosynthetic cells. Animals are, in essence, mutated plants. Many of the anaerobic bacteria, which die in the presence of oxygen, are the direct descendants of the prephotosynthetic cells. They survived in small out-of-the-way niches which still exist today.
The Animal MutationThe photosynthetic cells, once formed, must have quickly covered the surface of the entire ocean. In the process they began to absorb all the CO2 in the oceans. As this CO2 was absorbed, more of the atmospheric CO2 was dissolved in the ocean, and the total concentration of free CO2 began to decrease. At the same time there was a cooling of the earth from a quickly decreased greenhouse effect. But the sun was probably getting hotter.Simultaneously the photosynthetic cells were releasing oxygen into the oceans and the atmosphere. At first the oxygen was absorbed by the reduced minerals, particularly iron. The earth first had to rust before it had an oxygen-rich atmosphere. Eventually the atmospheric concentration of CO2 was so low and the concentration of oxygen and photosynthetic cells so high that these cells began to die, choking on their own wastes and starving from a lack of nutrients. Somewhere about this time, one to two billion years ago, occurred the mutation which was to lead to humanity.
The primitive photosynthetic cells still exist in the form of blue-green algae. Fossils (stromatolites) produced by blue-green algae 3.5 billion years ago in Australia were discovered in 1986. The green and red algae represent more highly evolved photosynthetic cells. Cells with properties of both photosynthesis and animals occur among the bacteria. Sometime after nutrients became scarce for the photosynthetic cells it became advantageous for some cells to metabolize the bodies of their dead relatives. This was an added ability which enabled them to profit from the deaths of the purely photosynthetic cells. Eventually these cells became highly mobile, like the amoeba, and they began to prey on the living algae. Once this started, since there was an enormous excess of algae, there was an advantage in changing their metabolism to strict oxidation of the bodies of algae and in stopping photosynthesis: i.e., they began biosynthesis. In the process these cells became oxygen breathers and lost their photosynthetic capability. Eventually a balance was established in which some cells metabolized through photosynthesis, some metabolized through biosynthesis, i.e., the oxidation of other cells, and some had properties of both. The former became plants, the latter became various types of bacteria and fungi, and those in between became animals. We define as "bacteria" all free living cells without a nucleus. Geneticist Lynn Margulis, although she did not use the term "autopoiesis," developed the now widely accepted theory that nucleated cells developed from autopoiesis within colonies of bacteria.
If there was a proliferation of plants, it would soon be counterbalanced by a proliferation of animals which would metabolize the plants and their by-products. If then the concentration of plants fell too low, there would be mass starvation among the animals, who had lost their capability for photosynthesis, and this would lead to a new proliferation of plants. The photosynthetic bacteria as well as the old anaerobic bacteria served as a buffering agent between the two newer life forms. However, the evolutionary pressure was such that the long-term advantage belonged to the animal cells and not to the algae or bacteria.
Sexual ReproductionAll the early cells reproduced asexually through binary fission in a process analogous to the splitting of the protocells. In binary fission (mitosis) the DNA inside a cell replicates itself, the two copies then segregate themselves at separate poles of the cells, then the cell splits into two identical copies. In sexual reproduction (meiosis), two different cells share their DNA and combine into new patterns after splitting into two haploid cells, each with half the DNA of the original. Each cell gives half of its DNA to the other and becomes a unique combination afterward which splits by mitosis.
Within the cell the DNA usually is not a single, paired molecular strand except in the most primitive cells and viruses. (We note that some viruses are based on RNA and not DNA.) Instead the DNA is organized into discrete, complementary paired packages called chromosomes. Each chromosome pair is somewhat like a book of redundant one-line instructions divided down the middle. Each half is necessary to complete the full message for each line. Although sterile haploid females can be produced from an unfertilized ovum among the lower species, no one has done this for the higher animals. In sexual reproduction each cell divides the books of its chromosome pairs in two and exchanges halves with another cell. If both cells were identical, the sexual reproduction would be no different from the asexual reproduction in its effects. However, no two cells are ever absolutely identical, since they each have had some mutations which would alter their DNA blueprint in different ways. Each mutation represents some change in one of the one-line instructions. Males also have one chromosome which is uniquely male; they share all their other chromosomes with females. Therefore, when the two halves of the different cells are recombined, they will form a completely new set of instructions in some lines and lead to a unique new individual.
Sexual reproduction has the de facto effect of increasing the mutation rate without significantly increasing the rate of deleterious mutations, since what is being done is combining two different sets of viable instructions into a new pattern. It so happens that some instructions which are evolutionary by themselves are entropic when combined. Rh-factor disease in humans is an example of this; so sometimes hybrids are inferior to either parent. But often the hybrids combine the best features of both parents and are superior to either parent. This phenomenon is referred to as "hybrid vigor."
The life function that any instruction controls is determined by its position on the chromosome, i.e., its position on a particular page of the book. The set of all chromosomes is the library of instruction manuals for regulating the biological processes of all living creatures. Each library is unique to each species and cannot recombine with the library of another species except through radical laboratory procedures involving recombinant DNA that produce interspecies hybrids called "genetic mosaics" [584]. Species differentiation occurs when two groups of the same species are isolated from one another so that they cannot interbreed and therefore undergo different mutations; their libraries, that is, their chromosomes, become so different that they can no longer recombine at random, because too many nonviable combinations occur or they are chemically incompatible.
In evolution there is a progression of integration by twos: (1) Some nucleic acids and proteins integrate into symbiotic systems of nucleic acids and proteins (the protocells); (2) some nucleic acids integrate into a symbiotic system of two-stranded DNA (primitive cells); (3) some DNA integrates into a system of paired packets of DNA, the chromosomes (advanced cells); (4) some species integrate into complementary pairs called "sexes." An extrapolation of this process is given later by which evolutionary hierarchies by systems of four complementary pairs are shown to exist at all levels, from atoms to super-human societies.
All the higher, i.e., least entropic, species reproduce by sexual means. Even some primitive bacteria exchange genetic material within their colonies, although this is not done in as obvious and seemingly purposeful a way as in the paramecium. Through sex the potential variability of a species is given its maximum expression, and the progress of natural selection works more effectively. Each individual in a sexual species must not only find nutrients and go through the physical motions of reproduction, it must also find and compete for a mate. The competition for and selection of mates is an important part of natural selection for all the higher animals, particularly humans, as Darwin first eloquently noted [97, 173, 245]. However, before discussing natural selection among humans, we will examine how it has functioned in all the animal species that led to humanity.
For now we note that each individual in a sexual species is far more different from its siblings than individuals in a nonsexual species. Indeed, the probability of the same parents producing genetically identical twins from two distinct sets of ova and sperm is less than 10-100 [744], which is close to impossible! This is why we are all unique. Identical twins come from a single, split embryo, not two separate genetic combinations. The higher degree of useful differentiation which occurs in a sexual species tends to produce more generalized capability within the species as a whole for predicting and controlling the total environment. Each individual represents a unique combination of properties which can serve as a base for one more step up the evolutionary ladder.
Animal EvolutionThe evolution of animals is of central concern in studying the evolution of humans because humans evolved from simpler animals and all animals evolved from the protozoa. However, the evolution of animal life is inextricably interwoven with the evolution of plant life. The plants are our partners in evolution - our complementary pairs in the biosphere.
The protozoa consist of all single-celled organisms which have no photosynthetic capability and depend on consuming products or entire cells of living organisms. The fossil evidence about the evolution of the protozoa is meager because in general they have no hard parts and leave no fossils. Certain bacteria secrete unique organic compounds. Deposits of these compounds have been found in rocks over 3.8 billion years old. Therefore we have evidence that the bacteria are about four billion years old. The protozoa evolved after the earth had an oxygen-rich atmosphere, about one-to-two billion years ago. The early protozoa were probably all plant eaters. As the protozoa expanded and plant food became scarcer, some protozoa began to prey on one another.
This is speculation, but remember many essential facts are missing. We are constantly speculating to complete the evolutionary picture, which itself evolves by trial and error. Everything in this book may be in error including all the "hard scientific facts."
Predation on one another by protozoa established a new balance in nature in which there were now four higher types of life forms: (1) plants, i.e., algae, (2) plant-eating protozoa, (3) protozoa-eating protozoa, and (4) various combinations of the first three. The fact that protozoa began to prey on the more active animal cells caused evolutionary pressure to favor mutations which enhanced defensive mechanisms among the prey cells and more effective sensory and mobility mechanisms among the predator cells. Some prey cells concentrated inorganic salts into a protective shell, e.g., foramenifera and radiolaria, and this produced the first true microfossils of about 2 billion years ago. The famous white cliffs of Dover were produced by these microorganisms, as were many other geological features such as the Mesabi iron deposits in Minnesota.
Another method for protecting themselves from predators was for the protozoa to organize into colonies which were too large for an individual predator to ingest. Some of the earliest microfossils, from 1.5 billion years ago, are of these colonial protozoa. Once an evolutionary advantage was established to colony formation, then the individual protozoa, under pressure of natural selection, began to organize themselves into ever larger and more complex interdependent societies which were to evolve into multicellular animals - the metazoa.
The MetazoaWe are very fortunate in being able to witness a living example by which separate, individual cells organize themselves into a complex interdependent colony. This occurs in bacterial colonies and the slime molds, which are formed by amoebalike creatures which combine features of protozoa and fungi [697].
The slime mold single cells live as independent amoeboid predators similarly to the regular amoeba. When some of the individual cells are ready to reproduce they secrete a chemical substance called "arcasin." This serves as a chemical attractant and stimulant to other individual cells which migrate toward the secreting cell and in turn secrete more arcasin, thereby attracting and stimulating other cells until a colonial mass of many thousand cells exists. This mass then functions as a single individual and crawls wormlike toward a suitable nesting site where it roots, grows a stalk, develops spores and casts them out to become new individual amoeboid predators which will repeat the cycle again when the conditions are right.
A much more dramatic example of metazoan evolution is contained in human embryology. Since all animals have evolved from single cells by a series of mutations in the basic DNA molecules, and many changes in the DNA molecule become part of the permanent record, the embryological development of a human or any other animal should mirror the entire evolutionary history of its species from a single cell. This is indeed the case at the embryological, not at the adult, level.
The fossil history derived from paleontological findings confirms the embryological evidence that the basic progression went from single cells to cell colonies, to interdependent systems of differentiated cells, to wormlike animals, to fish, to amphibians, to reptiles, to primitive opossum-like mammals, to generalized placental mammals, to tree-dwelling primates, to ground-dwelling primates, to successively more advanced types of hominids. The hominids are all upright-walking primates, which broke off from a common ancestor to both the chimpanzees and humans about five million years ago.
From the beginning of the earth until about 4 billion years ago, there is no fossil evidence, since this was the time of chemical evolution. From about 4 billion to about 2 billion years ago there were no higher life forms than photosynthetic cells. From 2 billion years ago, there is no evidence of any life higher than one-celled protozoa until one billion years ago. From 1 billion to 500 million years ago, there are nothing but growing quantities of invertebrate fossils: sponges, jellyfish, worms, mollusks, trilobites, echinoderms, sea scorpions, etc. With the possible exception of some worms, none of these life forms are in the ancestry of humans. From 500 million to 325 million years ago we see no higher vertebrate than the fish. From 325 million to 280 million years ago, the highest vertebrates are represented by the amphibians, then the reptiles appear and represent the dominant form of land life for over 200 million years. During this period the mammals appear, evolving from early mammal-like reptiles about 200 million years ago. The birds split off from the reptiles at about the same time. It takes mammals over a hundred million years to replace the reptiles as the dominant form of land life after the mass extinction of the dinosaurs 65 million years ago. The mammals reach their maximum diversity and numbers about 20 million years ago and then begin to undergo a gradual decline in number of species. At about the same time, the primate line that is to lead to humans, the Hominoidea, begins to diversify and proliferate. From the fossil evidence, humanlike apes appear about 18 million years ago, e.g., Sivapithecus and Ramapithecus. Apelike hominids appear about 4 to 5 million years ago. Our fully human ancestors, indistinguishable in any important biological feature from us, appear about 100,000 years ago in Africa and the Middle East. Extensive fossils have been found only from about 50,000 years ago, in Europe. In the last 100,000 years Homo sapiens has become the clearly dominant form of animal life and threatens, like all other previous radical mutations, to outbreed its food supply.
The Myth of Ecological BalanceThe history of evolution is the history of succeeding ecological catastrophes. The survival value of photosynthesis occurred after the first cells had just about exhausted their food supply and were facing mass starvation. The mutation which led to animal cells occurred after the algae had just about exhausted their food supply and were facing mass starvation. The individual prey cells mutated into metazoa when the more specialized predator animal cells began to put evolutionary pressure on the more generalized omniverous prey cells by consuming them in ever greater numbers and threatening them with extinction. Once the simple metazoa came into being, some of them in turn mutated into predators that preyed on other metazoa and the larger single cells. This established a new ecological cycle in nature. The existence of metazoa created a new ecological niche for the algae, bacteria, and protozoa in which they could exist as parasites on the larger, more evolved life forms. The one-celled life forms had in turn been hosts to a special type of parasite called a "virus."
A virus is a degenerate form of life which has lost the capacity to reproduce except within the living cell of a host. It has regressed toward the protocell stage and below. Contrary to some theories, viruses are probably not the ancestors of cells but are instead mutated degenerate descendants of cells or protocells which can only exist as parasites and are not independently self-replicating. A parasite is a life form which has higher entropy than its ancestors and can only live at the expense of a life form which has lower entropy. As we shall see, the induction of parasitism is an ecological imbalance which is an intrinsic part of the evolutionary process and occurs even among the highest species, the humans.
It is the very imbalances in the ecological system which provide new opportunities for evolving life forms. The appearance of land plants about 400 million years ago created an ecological imbalance which made it possible for land animals to evolve from the fish and arthropods. The early amphibians created an ecological imbalance by filling a new ecological niche and were almost all quickly replaced by their descendants, the reptiles. Of the wide variety of amphibian life which flourished 250 million years ago, only the frogs, newts, and the salamanders survive. The reptiles flourished for almost 200 million years and with catastrophic suddenness began to die out.
There is considerable evidence that approximately every 26 million years there are catastrophes of cosmic origin, e.g., collisions with comets, which wipe out many of the specialized species and open new niches for the most intelligent generalized species. One of these catastrophes apparently occurred 65 million years ago when the dinosaurs were wiped out and replaced by the mammals. Those periodic catastrophes appear to be due to a swarm of comets which intercept the orbit of the earth every 26 million years. There is a danger of the entire earth being destroyed at each interception if several very large comets hit it in quick succession.
Today the ecological imbalances that humanity is creating are primarily the effects of switching food supplies from serving the reproductive purposes of other animals to serving the purpose of providing for ever more human beings. The percentage of the total biomass taken up by any species is a biological measure of its evolutionary success. However, it is theoretically possible to destroy the basic cycle of life through pollution and other means [280]. It also appears that the human species produces at least as many parasites as it produces hosts for them. This is entropic.
It is usually sentimentality that makes some environmentalists decry the extinction of tigers, wolves, and other large predators which have almost nothing to contribute to human evolution and are incapable of competing with the prime predator, Homo sapiens. But it is legitimate to decry the total sterilization of large parts of the ocean or destruction of the ozone layer and more through pollution [280]. We note that the trend in human evolution seems to be to domesticate all life forms, including ourselves, which serve human purpose, and to replace the rest. If ecological balance could ever be achieved, it would probably mean the end of evolution.
The Tree of LifeThe Tree of Life, illustrated below, represents the approximate kinship of the various life forms and shows their departure from the main trunk of evolution as a function of time. Traditionally humanity has been shown at the top of the evolutionary tree. This was done more for emotional, egocentric reasons than for any more fundamental underlying criterion. However, we will show that this is where humanity belongs.
A common criterion which has been used by some evolutionists as determining the level of evolution of a given life form is its degree of change from a more primitive ancestor. By this criterion, elephants may appear more evolved than humans, in terms of their exterior morphology, if we trace the degree of change from a common ancestor. Intuitively, we feel that this may be wrong.
Change by itself does not determine evolution, since a change may increase entropy. This is obvious in the entropic decay of parasites which are very different from their ancestors. We defined an evolutionary improvement as a change which decreases entropy and increases the complexity of the organism. But it is not obvious that a human is more complex than an elephant. An elephant has a larger body and a larger brain than a human. Still we may intuitively feel that the elephant is less complex than a human. --The Tree of Life--
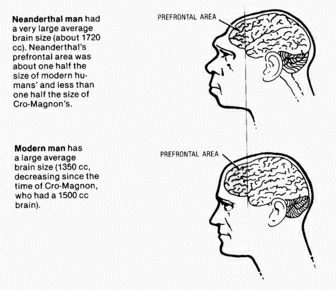
Without getting into neuroanatomy, we can say that although the elephant brain may be larger than a human's, it has a simpler structure and therefore is a simpler brain. This is immediately obvious in terms of the behavior of elephants, which can in no way approach the complexity of human behavior. Since behavior is well known to be regulated by the brain, we must assume that the human brain must be more complex than the elephant's.
In discussing the evolution of humanity's immediate ancestors, we will go into more detail about the relative importance of various brain subsystems; for now we merely assume that there is a one-to-one correspondence between complexity of brain and complexity of behavior. In general, larger brains are more complex than smaller brains, but not always, since a large part of the brain may be used for controlling the vegetative routine functions of the animal. If we consider brain size in relationship to body weight, we get the relationships shown in the following table:
Ratio of Brain Weight to Body Weight
in Selected Animals (adapted from Tobias [777])
Mammals
Squirrel monkey (Saimiri sciutea)
Tamarin (Leontocebus)
Porpoise (Dolphin) (Phocaena communis)
House mouse (Mus musculus)
Tree shrew (Tupaia javanica)
Man (Homo sapiens)
Ground shrew (Sorex minutus)
Monkey (Macaca mulatta)
Gorilla (Gorilla gorilla)
Elephant (Elephas indicus)
Sperm whale (Physeter catodon)
Reptiles
Crocodile
Stegosaurus
Brontosaurus (Apatosaurus)
Ratio
1:12
1:19
1:38
1:40
1:40
1:45
1:50
1:170
1:200
1:600
1:10,000
1:5,000
1:30,000
1:100,000
We note that humans are considerably better off than elephants but not better off than a mouse [777]. This is explained by the fact that complex interactive behavior, e.g., learning abilities, is controlled primarily by the neocortex and not by the rest of the brain, which controls the vegetative and automatic responses of the body such as the metabolic cycles and emotions. If we plot complexity of behavior as a function of the fraction of brain weight which is taken up by the cortex, then we see the type of direct correspondence that we would intuitively expect.
For the time being, we will assume that (1) the major direction of evolution is toward the development of ever more complex behavior as a means of increasing reproductive effectiveness; (2) complexity of behavior is in one-to-one correspondence with the complexity of the nervous system; and (3) a rough approximation to the complexity of the nervous system can be made by multiplying the fraction of the nervous system taken up by the cortex against the mass of the nervous system to determine "excess" neurons, as a crude surrogate measure of this complexity. From a different perspective we can say that the brain is a self-organizing, biological computer. The cortical parts, particularly the neocortex, are a general-purpose computer which are easily programmable and above all self-organizing.
As we shall later see, reality is actually much more complex than this. But this gives us a first crude approximation for discussing the direction of evolution and the meaning of the tree of life.
Direction and MeaningIf we again consider the Tree of Life, we see that it is continuously sending out branches from the main trunk. These branches in turn put out many twigs, but eventually stop growing and often die. Only the main trunk continues to grow upward. Once any life form branches off from the main trunk, it only moves ever further away from the trunk and it never rejoins it. Eventually all branches must stop growing, i.e., all species which branch out will eventually become extinct and leave no progeny.
The major differences between the species which are along the main trunk and those which are along the branches is that the former are generalized while the latter are specialized.
A generalized species is one which may obtain nutrients from many different sources and which may survive in many different environments, although it is less efficient in doing so in any particular environment than a "specialist" in that environment. A specialized species is one which may obtain nutrients very efficiently from only a few sources, but can survive only in a narrow range of environments. When a species specializes it is on its way to extinction because even slight perturbations in its food supply or environment can cause all its members to die out. The periodic extinctions of every 26 million years keep eliminating the specialized species, making more room for the most intelligent generalized species. This is equivalent to a "pruning" of the tree of life. Remember, this is speculation.
Once evolutionary pressure begins to pull a species toward specialization, it can only become ever more specialized along the direction it has chosen. Through mutations it gains characteristics which enhance the specialization and eventually loses characteristics which are no longer relevant. Mutations driving the species backward toward the trunk are less likely to occur and are less likely to survive than mutations driving it out toward the end of the branch. Therefore, the species begins to become a closed system existing within an ever narrower environmental pattern until entropy destroys it.
It should be noted that under natural selection specialization is inevitable. If there are ecological niches which specialized species can fill, they will mutate from a more generalized species until these niches are filled. The higher any species is on the evolutionary ladder when it begins to specialize, the more effective it will be within each specialized niche. However, a generalized species does not begin to compete for a specialized niche unless it is vacant. This is what happened when the specialized dinosaurs suddenly became extinct about 65 million years ago. The generalized mammals and some birds specialized and filled all these niches within 26 million years. Then they too began to undergo extinction. The generalized species are usually at a temporary disadvantage relative to some more specialized species; therefore they are usually relatively small in biomass at a particular point in time, but they are larger in biomass over a long period of time. An example of this is the opossum, a small, highly generalized, primitive mammal which can exist in a wide variety of environments. The opossum, although never great in total biological mass if compared to other more specialized animals, has existed in its present form for millions of years. Furthermore, the opossum is less complex than many placental mammals, such as the horse, which underwent extinction in the environment it shared with the opossum.
Therefore, although the direction of evolution is toward ever greater complexity, complexity by itself does not assure survival and almost certainly guarantees extinction if it is a specialized complexity. The direction of evolution is toward ever greater generalized complexity. The lesson of evolution is that specialization leads only to extinction. Generalization is a necessary but not a sufficient condition for a species to continue its climb up the evolutionary ladder.
The Top RungAt any given time the top rung of the evolutionary ladder is occupied by the most complex generalized class of animals. About 65 million years ago this rung was occupied by the mammals. As the mammals began to specialize, to fill the niches of the extinct dinosaurs, although they increased their complexity for a while, they eventually climbed down the ladder, increased their entropy and many fell off, i.e., became extinct. About 40 million years ago the top rung was occupied only by a single generalized group, the primates.
The primates were highly generalized in the beginning and probably had an appearance very similar to today's tree shrew. Like all generalized groups, the primates began to shoot off specialized branches. The early primates were probably equally adept at living in trees or on the ground, but were not outstanding in any one of these modes of life. They all were omniverous and had grasping hands and feet which they used to good advantage for getting food and tree climbing. As the primates began to specialize about 40 million years ago, they began to emphasize one way of life over another - land dwelling or tree dwelling. In the process they began to become more specialized.
For living on the ground there is considerable advantage to running on all fours. Therefore, one family of primates became mostly quadripedal, as are the baboons and mandrils. For living in the trees there is considerable advantage to having long powerful arms for swinging from branch to branch (brachiating); therefore, another family of primates became mostly arboreal with long powerful arms and light body, as are the various groups of monkeys (rhesus, macaques, capuchin, etc.). Another group remained generalized in between, not fitting into any of the environments as well as its more specialized cousins. This group is represented today in varying degrees by the anthropoid apes, also called "hominoidea," e.g., gibbons, orangutans, gorillas, chimpanzees, and humans. They began to differentiate from other primates about 20 million years ago.
For millions of years the hominoids made up a very small percentage of the primate biomass because they could not compete effectively against their more specialized cousins. This is shown by the paucity of hominoids fossils, which are very scarce up to about 5 million years ago. (Part of the problem is that tree-living species rarely became fossilized.) But as we have seen, the evolutionary future belonged to the hominoids because once the other primates had specialized only the hominoids occupied the top rung of the evolutionary ladder.
Hominoid EvolutionSome of the hominoids continued along the main trunk of evolution up to the present time. About 18 million years ago a series of mutations occurred which enabled one group to become primarily land dwellers without becoming less generalized. This was made possible by substituting some partial, bipedal ground-running capability for tree-climbing ability and by acquiring at the same time a new capability of better use of the hands whilein motion. The net effect was probably a slight increase in overall general capability. At first this probably gave little or no advantage to these animals, some of whom were to be our ancestors, but what it did was open a new ecological niche for the primates - the niche of a machine-using species.
Machine BuildingMany species have been and still are machine builders. For example, wasps, ants, and bees build complex nests which represent a community machine. Beavers build complex systems of ecological control with their dams, canals, and partially submerged houses - minor Venices. Beavers must gather branches, make mortar, dig canals, cut down trees, and arrange everything in a special order at the right places and keep all their machines in constant repair.
A machine is a manufactured device which transforms one form of energy into another. Tools, language, and social organization are all examples of machines. A machine decreases entropy for an animal which properly uses it by increasing the amount of useful energy at its disposal. A machine-using animal can, as a consequence, enhance its ability to acquire nutrients and reproduce beyond those abilities of a comparable animal which does not have a machine to use.
Many animals build and use machines. The capability for doing this, i.e., the blueprint for the machine and the operating manual, is transmitted by the genes and structured into the nervous system. The blueprint is "hard-wired". This is clearly shown in experiments which demonstrate that the nest-building capabilities of insects, birds, and to a lesser extent beavers are not learned but are a product of inherited instinct [470]. The machines of insects and birds have remained the same for many millions of years [93]. They have never acquired the capability to build any machines other than nests. The instinct for machine building is highly specialized and not generalized.
The beavers, on the other hand, have shown some changes in their nest structure over the last few million years. Their machine-building capabilities are only partially instinctive (hard-wired), although this is the main part. The non-instinctive part, which is more generalized (soft-wired), enables them to make slight variations in design. This is a result of the fact that they have much more complex nervous systems than both insects and birds. However, the beavers are already highly specialized for an amphibious way of life and their bodies are adapted to building only a special type of machine - an ecological community. They do not really create new machines but only make slight variations on an existing model which they carry in their genes. Our hominoid ancestors were different.
The early generalized hominoids which had slightly better bipedal capabilities than their more arboreal cousins, had no instincts for building machines. These hominoids, which are called "hominids," only had the advantage of being able to have their hands free while walking on the ground. They were not particularly good runners, compared to the baboons, for example, or as good climbers as their immediate cousins, the pongids, who were to evolve into the orangutan, gorilla, and chimpanzee. If they had not made maximum use of their hands almost from the beginning, they probably would have become extinct. The mutation for bipedal walking would have had no net survival value by itself.
The current chimpanzees can create machines, albeit very simple ones, independently of instinct. A chimpanzee will pick up a fallen branch and use it as a club. This, however, is using a natural object as a machine, not creating one. There is no real manufacturing involved. In their natural state chimpanzees have been observed to take a small branch and purposefully strip off all twigs for the express purpose of using it to capture termites [794]. This is not done by instinct; rather, it is either learned from another chimpanzee, or it is independently discovered through the act of experimentation.
The fossil evidence indicates that the immediate ancestors of the hominids had nervous systems comparable to a modern-day chimpanzee's. Therefore, they probably had at least as much machine-making capability. From this it follows that the early hominids probably used natural branches and stones as simple tools and could probably make simple but purposeful modifications and choices such as stripping a branch of all twigs or choosing a conveniently shaped stone for a specific purpose. This is enough to have given a slight advantage and opened a new way of life to our ancestors who acquired a bipedal capability.
Hominid EvolutionThe evolution of humanity has been the evolution of the capability to create and use machines. Once the hands were free while walking, even if this was only a temporary freedom, there was a strong evolutionary pressure to enhance bipedal walking capability. The greater percentage of time the hominid could remain upright and keep his hands free, the better off he was. By about five million years ago, the hominids had acquired the upright posture characteristic of modern humans and their hands and arms were almost entirely devoted to making, carrying, and using machines while on the ground. This is also the period when chimpanzees and hominids separated from a common ancestor. The gorillas are generally believed to have separated about ten million years ago, the orangutan about 14 million years ago, and the gibbons about 20 million years ago. This is confirmed by DNA dating [600], a method for determining how close species are genetically in terms of when they last shared DNA with a common ancestor. Some very recent evidence not yet published by anthropologist Vincent Sarich and his associates indicates that hominids, chimpanzees, and gorillas all separated from a common ancestor about five million years ago. However, the previous figures are more commonly accepted.
In the case of the orangutan, gorilla, and the chimpanzee it is likely that their immediate predecessors were more like hominids than like them, e.g., Sivapithecus and Ramapithecus. The knuckle-walkers (pongids) may represent early specialized hominids. From about five million years ago, the hominids were clearly inferior to the pongids in tree-climbing ability, but they were far better walkers and runners. From the neck down the hominids were structurally almost indistinguishable from modern humans except that they were shorter and more rugged.
At the same time that the hominids were changing from the neck down, another change was occurring in the skull. Among the first changes was probably the loss of the great canines which characterized the pongids. These canines are used primarily for fighting among the male pongids for the purpose of establishing dominance and having first access to the females and food. It is a characteristic of the evolutionary process that any organ or structure which plays little or no direct role in survival will atrophy, since there will be no natural selection against the mutations which cause it to atrophy. Once the hominids began to depend on tools for fighting and gathering food, there was a clear survival advantage for the more efficient tool users over those who depended more on their large canines. A tool is a machine which transforms energy from the muscles, bones, or teeth directly into useful work. It might be considered an amplifier of our body structures. Therefore, the canines atrophied and the shape of the mouth and face changed. This eventually made modern language possible. However, there was relatively little development in brain complexity during this time, which was also essential for language.
So long as the hominids were filling a new ecological niche somewhere between the baboons and pongids they merely expanded over the parts of the earth which were hospitable and which they could reach. However, it is likely that all major evolutionary progress of the hominids from earliest Australopithecus to Homo sapiens occurred in Africa. There is apparently something about Africa that is conducive to hominid evolution - perhaps its ecological richness.
The main pressure was on developing full upright posture and tool-using capability. Stone-throwing gave hominids a unique biological advantage and contributed to the development of the nervous system in determining trajectories and in choosing and handling the right kinds of stones. The early hominid expansion was limited to the temperate zones of Africa. The early hominids were still dependent on the trees for safety, but they could wander further away than pongids because of their running and tool-carrying capability. There was, at this time, little evolutionary pressure to enhance their tool-making skills because there was a large part of the world to be filled and the hominids had little competition from other primate tool users. From very humble beginnings, the hominids would expand and fill the ecological niches of a tool-using species which got most of its food from the ground but still was partially dependent on trees for safety and some food. Once these ecological niches were filled, then the hominids began to compete against one another and further develop the brain. (Remember, we are speculating.)
What happened was that because the hominids lived in widely scattered, relatively isolated small bands, considerable evolutionary development could occur in one hominid group without affecting another. At this time the total hominid population on the earth, virtually all in Africa, was probably not more than a few hundred thousand. There are about three hundred thousand chimpanzees today, all native to Africa. The chimpanzees have remained virtually unchanged during this period. Once one group of hominids had a clear advantage in its machines and/or their use over other hominids, it would begin to re-expand into the territory of its less developed cousins. Through natural selection the hominids with better machine-making capability and/or social organization would replace the less evolved hominids.
Human social organization is a machine in which the components are human beings. Therefore, hominid evolution became primarily one of internal competition between different hominid groups. This does not mean that there was a purposeful genocide by one group against the others, but merely that the more evolved groups preempted the food supplies and out-reproduced the less evolved groups. Once this process started, about five million years ago, there was considerable pressure put on brain evolution for the purpose of creating more complex machines, including hominid organization. During the last two-and-a-half million years the average hominid brain was to triple in size.
One of the consequences of developing an upright posture was a modification in the hominid pelvis which severely restricted the size of the birth canal. This meant that expansion of the brain was only possible if the young were born at ever more immature stages of development. Therefore, the young were completely dependent and highly vulnerable for a long time after birth. This produced evolutionary pressure for the development of emotional programming in the brain that would be conducive to the formation of family units in which the mothers and their offspring were protected and provided for by the males. The process of increasing immaturity at birth began once upright posture began to have clear evolutionary advantage and continued until about 50,000 years ago, at which time the disadvantages of extreme immaturity at birth outweighed the advantages of greater brain size, primarily because of an excessively high infant mortality rate. At the same time the interdependence between male and female became even more pronounced.
The extreme immaturity and long-term dependency of the hominid newborn made it imperative that the young be under constant maternal care for many years after birth. It was very difficult for a hominid female to gather food, let alone hunt, while caring for the young or in the latter stages of pregnancy. Therefore, the hominid males had to become emotionally predisposed to provide for and protect the females with whom they mated, or the hominid line would have become extinct. At the same time the female had to become attached to a male protector once she become pregnant. This mechanism is known as "the sexual bond." It occurs among many mammalian and avian species, particularly predators. It was probably enhanced among the hominids by the relatively higher sexuality of the females. Today the only female animals that are known to experience orgasm and are almost always sexually receptive to their mates are humans. The long-term nurturing instincts of the female, together with the growing socialization and protectiveness of the males, made it possible for the early hominids to form into mutually supportive family bands.
To a certain extent this banding into family units exists among all primates. The baboons, which share with the early hominids the dangers of predators when they are away from trees, are organized into bands called troops ranging usually from 15 to 50 individuals. The females and young stay near the center of the troop and are protected by the adult males who guard the periphery of the troop. The males themselves are organized into a rather complex hierarchy of dominant groups and dominant individuals within each group. If a member of a male group is threatened by an outsider seeking to assert his dominance, the whole group will cooperate as a unit to ward off this threat [220]. This hierarchy of structure imposes a group discipline which permits the troops to have social stability and to function together as a unit, very much the way a military organization does. The evolution of baboon social behavior required many millions of years of natural selection which centered primarily on changes in the emotional centers of the brain, as opposed to the reasoning centers in the neocortex which are used in machine building and cognitive processes.
The early hominids were probably more like the chimpanzees, who most closely resemble the protohominids physically and are closest in their genetic and emotional makeup to humans, as opposed to the more specialized baboons. The chimpanzees do not have such a rigid or complex hierarchical social structure as the baboons. A baboon may stay in the same troop it is born into all its life. Chimpanzees move from one family group to another with considerable ease. The male-dominance pattern of chimpanzees is more individualistic and does not involve as much organization into aristocratic cliques as among the baboons. The females live closer to the trees and are readily able to provide for and protect their young without help from anyone. Yet humanity's current social behavior is much more like that of the baboons than the chimpanzees.
As the hominids were spreading over the earth they were at first probably organized into small family units of one dominant male and probably not more than four adult females and their young. A bonding mechanism evolved such that the male assumed responsibility for each female with whom he mated and her young, and the female became faithfully attached to and dependent on the male. This is the "evolutionary contract" within our species. This type of social structure occurs currently among the gorillas, who are in some ways more like the early hominids than the chimpanzees, since they spend a larger percentage of their time on the ground, and only the young and the females climb trees for protection. The difference is that the gorillas have become specialized to an exclusively vegetarian diet. The male gorilla, who has difficulty climbing trees, protects the much smaller and more vulnerable females and young by warding off almost all predators (as opposed to scrambling for a tree) by virtue of his extreme size and strength. The females with young are not nutritionally dependent on the male.
The gorilla is more specialized than the chimpanzee in terms of diet and body structure. This type of specialization, together with human predation, is slowly bringing about the extinction of the gorilla, since its niche overlaps both that of humans and the chimpanzee, each of which is more adept in its sphere. There are only about 15,000 gorillas left in the world as opposed to several hundred thousand chimpanzees and five billion human beings. However, the large increase in the number of human beings is mainly due to a psychosocial phenomenon of the last 50,000 years in general and the last 10,000 years in particular (which will be discussed later). Up to about 50,000 years ago, the hominid population of the earth was less than one million. For now we can assume that the early hominids had an emotional structure somewhere between a chimpanzee's and a gorilla's and were evolving toward a social structure similar to a baboon's.
The early hominid families would probably cast out the male young as they matured, and the female young left when they mated outside of the band. The male sought to have as many females as he could protect and provide for and avoid competition from any other male. The early hominids were not primarily hunters but gatherers and probably scavengers, driving hyenas and other smaller scavengers away from the kills the large predators left behind, probably by throwing stones. As they adapted better to a walking and running existence, they began to include more meat in their diet, and eventually they became primarily hunters.
During the first wave of hominid expansion, which lasted about three million years and was limited to Africa, the hominids' hunting was probably limited to capturing the young and lame of other species, including the hominoids and possibly an occasional hominid from another group; so the chimpanzees hunt today. Eventually the hominid dexterity in the use of clubs and stones was sufficient to start bringing down more difficult but still small game. Today chimpanzees throw stones at potential predators to ward them off, but they can not bring down game in this way, at least not reliably. During this phase of evolution hominids began the first fierce competition amongst themselves.
Just as the hominids were acquiring full upright posture and filling all ecological niches available to them, climatic changes were causing the vast tropical forests which covered much of the earth to be replaced to a great extent by grasslands. This was most disadvantageous to the tree-dwelling primates, but it gave the hominids a considerable advantage. The upright posture of the hominids enabled them to see predators from a distance in the high grasses. The hunting skills they had acquired enabled them to do without the shelter and the food of the trees much better than the other primates. At this point the hominids, which had a precarious foothold on their niches, began to expand and all the other primates to decline. The tree-dwelling monkeys were later able to expand again into forests of Asia and South America, but the pongids were never to catch up to the hominids again, and some, such as the gorilla and the orangutan, are on the verge of extinction, primarily from human pressures.
Those noncortical mutations in the brain which predispose the hunting animal toward aggression and the cortical mutations which predispose him toward learning now began to be strongly selected among the hominids. About three-and-a-half million years ago a strain of hominids known as Australopithecus afarensis had a brain not much different in complexity from that of a pongid. However, Australopithecus was a fully upright omnivore and a hunter. He used tools of stone to help him catch and eat meat and to defend himself. Although he weighed less than 100 pounds and could not run as effectively as any of the large quadrupeds of the grassy African savannahs, he competed against all the predators and scavengers which abounded there. And in the end he won the battle. Early, not late, Australopithecus was almost certainly the ancestor of the modern humans.
The AustralopithecinesThe Australopithecines were a wide-ranging and varied group of hominids who began to spread over Africa about five million years ago. The rapidly receding forests opened new niches for them and they became a significant part of the primate biomass. They now exclusively occupied the top rung of the evolutionary ladder. Like all other generalized species at the main trunk of evolution, some of them began to specialize and fill in other niches. Australopithecus robustus, also known as Paranthropus, became a relatively large-bodied vegetarian who began to occupy the niche which had been left by the larger pongids. His large, flat rear teeth indicate that he might have given up hunting to concentrate on an exclusively vegetarian diet. He became extinct about one million years ago.
The oldest fossils of the genus homo (Homo habilis) seem to be about two-and-a-half million years old. The most recent skull of Homo habilis is from 1.8 million years ago (as of 1988). Therefore, the genus Homo coexisted in Africa along with the more specialized later Australopithecines for at least 1.4 million years.
It is clear that many of the early Australopithecines specialized into what are now known to be extinct lines of hominid development. The major form of specialization for the hominoids seems to be in the direction of becoming strict vegetarians. Those that choose this specialization become very large like the modern gorilla and the extinct giant hominoid Gigantopithecus. Part of the reason for increase in size is due to the necessity of having extra-long intestines for extracting as much food as possible from the less nutritious vegetation, which must be consumed in huge quantities. Gorillas, for example, spend an average of eight hours a day just eating. There is some evidence that the giant hominids (Meganthropus and his successors) which lived in southeast Asia until fairly recent times also became strict vegetarians.
Therefore, the main direction of Australopithecine evolution was toward a highly generalized hunter omnivore. So long as the Australopithecines were filling the evolutionary niches of a tool-using hunter omnivore in the expanding environment of the grassy savannahs, they were able to maintain their social structure of small family units with one adult male. At the same time, competition from other predators placed maximum evolutionary pressure on increasing brain complexity in order to develop better hunting skills - primarily by developing and more effectively using ever more complex tools. Eventually Australopithecus evolved into a large-brained predator, quite adept at solitary hunting. During this period, domesticated fire, shaped stone tools and probably spears were developed. Primitive spears were probably made by sharpening wooden poles on stones and hardening the points with fire. Shafted stone spears are a much later development. This enabled the solitary hunter to capture fairly large game, such as bushbuck, and defend himself against all but the largest predators. However, a large amount of the game biomass was in the large vegetarians such as the elephants, bisons, rhinoceri, etc., which were beyond the hunting capability of the solitary hunter and most of the large predators for that matter. Even a powerful predator such as a tiger cannot kill a full-grown, healthy elephant. As the small- and medium-size game were depleted by Australopithecus and other predators, the large game proliferated. Natural selection now favored the kinds of mutations which would lead to the formation of larger hominid hunting groups in a new type of social organization.
Hunting and EvolutionThe early Australopithecines would pass on their acquired hunting and tool-making skills to the young through example. Imitative behavior is a characteristic of hominoids in general and hominids in particular. Those Australopithecines that had the best brains would become the best hunters by virtue of having learned the necessary skills well and in rare cases by inventing new skills of their own. The best hunters could also attract and provide for the largest number of females and would, therefore, produce the greatest number of offspring. This provided the basic natural selection pressure for brain development. The young adult males would probably be driven out of the band when they began competing against their fathers for the females, or they would drive their fathers out if the old man had significantly weakened from age before the conflict arose. Therefore, only the most competent fighters and hunters reproduced. The human beard probably evolved during this period as a means of protection for the throat of the males in their mutual competition.
This same basic pattern exists among the gorillas. The adult gorilla is very tolerant and even protective toward immature males, but becomes much less tolerant after they mature. Similarly, adult male baboons behave protectively toward immature males but become increasingly intolerant toward them as they mature, unless they become totally submissive.
Natural selection worked on the female side primarily by her ability to nurture and care for her offspring and willingness to become bonded to a protective male. Therefore, natural selection worked on the emotional centers of the Australopithecine brain to produce maternal, submissive females attracted to dominant, aggressive males who were protective toward the females and their young but intolerant toward other adult males. The development of machine-making ability could not, of course, be strictly confined to one sex, since it is dependent on cortical structures. Only the emotional centers in the depths of the brain which are strongly influenced by the sex hormones, particularly during embryological development, could become highly sex-specific. Therefore, both males and females probably created machines, although not necessarily the same machines. However, the opening niche for hunters that could pool resources and capture large game put evolutionary pressure for further emotional changes in the male.
Group hunting would not have been an evolutionary advantage for the early Australopithecines because they did not yet have the necessary tools nor hunting skills for bringing down big game. Natural selection was based on the individual ability of each male and female. But about two million years ago, and possibly much earlier, group hunting was made evolutionarily practical by (1) the changes in ecology which Australopithecus had himself helped bring about by contributing to the depletion of small-and medium-size game, by (2) the proliferation of large game and predators, and by (3) his own increase in numbers and hunting technology.
In order to have group hunting, the males not only had to have tolerance toward other males, but they had to actively cooperate and share the kill with them. The mutation that made this possible was actually a rather simple one. It was merely necessary for the adult male to extend his protective, friendly attitude toward his adult as well as his immature children and for the young adults not to be sexually attracted toward their father's mates or their sisters. A certain amount of this type of emotional programming occurs among the pongids, which have been observed to not be incestuous, at least not between mother and son. Some experiments done with human children indicate that children who are raised in the same family, independently of their blood relationship, are not strongly attracted to one another sexually. This emotional programming against incest has clear evolutionary value, since it minimizes the possibility of combining deleterious genes and maximizes genetic variability. Therefore, it was not necessary for the adult male to drive out his male children in order to form an exogamous breeding pattern among primitive, preverbal hominid societies. We note that exogamous mating maximizes the variability of any species and decreases the probability of deleterious genes combining. It increases the rate of evolution as does sexual reproduction.
The emotional mutation which made adult males tolerant and protective toward their adult children probably also made them less aggressive toward other males, but not necessarily tolerant. Therefore, group hunting probably began as a family enterprise where the dominant male and his older sons cooperated to bring down game. This opened a new ecological niche for the hominids since now virtually all animals were at their mercy. The fossil finds indicate that hominids of 300,000 years ago were already killing and consuming large numbers of elephants and rhinos. This took the cooperation of many adult males. The early group hunting was almost certainly limited to smaller game.
As group hunting developed, the needs for exogamy and group cooperation would bring different families together for brief periods, if for no other reason than the exchanging of mates. This type of social structure currently exists among the Australian aborigines. When the proper occasions arose, a group hunt would occur in which two families would cooperate. The families were closely related through their matings and were akin to a clan. Eventually several families working together saw that they could capture the largest game. Groups of families numbering possibly up to two dozen individuals would then form into close-knit hunting units. Some of the North American Indians had this type of social structure. But the hominids who engaged in this large-scale group hunting were not Australopithecines; they were our immediate ancestors and belonged to the genus Homo.
The Genus HomoThe genus Homo differs from the most advanced Australopithecine primarily in having (1) a larger brain, (2) a larger size indistinguishable from modern humans, and (3) smaller jaws and face. The Australopithecines seem to have had a brain no larger than 700 cc. The brain of Homo erectus, who followed Homo habilis, averaged about 900 cc. A fossil of Homo erectus 1.6 million years old was found in Africa in 1985. However, it is not until the period of about 500,000 years ago that extensive fossil representatives of the genus Homo are found all over the world except Oceania, Antartica, and the Americas. The earliest evidence of large-scale group hunting is from less than 400,000 years ago [152, 96].
There may have been an ecological balance for over one million years between group hunters of the genus Homo, who were in the minority, and solitary hunting and vegetarian groups of Australopithecus. It is also possible, but not likely, that the mutation to the genus Homo was made several times independently in different parts of the world and that this led to the four major races of today. This is the theory of race formation presented by C.S. Coon in The Origin of Races [156]. It seems unlikely, but not impossible, in the light of other examples of parallel evolution. There may also have been some disadvantage to the mutation to the genus Homo which made these isolated mutations extinct - for example, intolerance by the much more numerous Australopithecines. In any case, there is no fossil evidence at this time of the clear dominance within the hominid group by Homo habilis until about 2.5 million years ago. Homo habilis evolved into Homo erectus in less than one million years. All this time Australopithecines of the later specialized varieties abounded on the continent of Africa. The exact mechanism by which the Australopithecines were replaced by the genus Homo can only be conjectured. What is known is that over a period of 1.5 million years, families of Australopithecines were completely replaced by group hunters of the genus Homo.
Homo erectus had a brain that was already within the size range of modern humans (900 to 1200 cc). He could kill any land animal through group hunting. His social structure may have ranged from the single family to a small clan of about eight families totaling about 32 individual persons. The major evolutionary pressure was on skill for inventing, making and using better tools and on a new machine - social organization.
Early Social OrganizationJust as a complex machine can be made through instinct, as in the case of social insects, birds, and beavers, so can complex social organizations be formed through instinct, as in the case of bees and baboons. Note that thesocial organization is a machine in which the components are living beings. The early hominid social organizations, such as the small and large families, were based primarily on instinct. But when groups of families began to form into larger, permanent hunting groups, there had to be social organization which was not entirely instinctive. A system similar to that of the baboons had to develop, which gave a hierarchical structure to the group and a chain of command. Baboon social organization developed through many millions of years of evolution. Group hunting developed among hominids in no more than two million years and possibly in as little as 500,000 years. This may not be enough time to develop the complex instinctive responses characteristic of the baboons. Furthermore, there is no trace of instinctive social organization in humans other than the family. What exists is the primitive primate characteristics of dominance and submissiveness.
The basic mutations in instinct which made group hunting possible were (1) the extension of dominant male tolerance toward other adult males and (2) the sexual avoidance of taboo females by the adult males. This together with hierarchical dominance is enough to establish a chain of command and to make it possible for many adult males to live together, but it is not enough to make them hunt as a coordinated group.
The hunting of elephants involved separating the prey from the herd and then killing it. Elephants are intelligent, highly gregarious, and protective toward one another. The fossil evidence indicates that some of the elephants were trapped in pits or stampeded over cliffs. This took planning, foresight, and pre-arranged strategy. This cannot be done without some kind of language.
LanguageSome anthropologists and linguists claim that humans could never have been group hunters without language. However, this is not necessarily true, as the group hunting of lions, wolves, and even chimpanzees indicates. So long as Homo erectus was hunting medium-sized prey, he could have probably gotten along without language by deploying around a prey and all attacking en masse on signal from a leader. But once this type of hunting began, there would have been considerable advantage to developing signs and signals for coordinating attacks and changing strategies when unusual situations arose. If this occurred, then there would have been evolutionary pressure to improve the communication system, and natural selection would have assured the survival and continued evolution of those hunting groups which had the best communication systems. So with the start of group hunting, the evolution of language was greatly accelerated.
We note that a rudimentary type of linguistic ability exists among the chimpanzees, who can learn, but not create, simple sign language. They also have clearly differentiated grunts for communicating.
The development of human language involved another modification in the neocortex and is probably the main cause for the rapid increase in brain size during this period. This represents a rate of increase in complexity unparalleled in evolution. There must have been an extremely strong evolutionary advantage for large brains, which began almost all of a sudden. The only feasible explanation seems to be the development of language, for the actual tools, although improved in this same period, do not represent that great an increase in complexity [152, 596].
Linguistic ability came into being long before there existed anything approximating a modern language. Homo erectus almost certainly used language primarily as means of communicating commands and storing simple facts, not for communicating the abstract thoughts which modern language is capable of [449]. However, this rudimentary language was a new machine destined for significant evolution. It made complex organization possible.
Through language it was eventually possible to tell someone to perform not merely a single set pattern, but a whole series of contingent actions. Organized hunting of the most complex kind was possible and likewise the corresponding social structure with complex hierarchies, duties, taboos, etc. The evolutionary pressure for tool building was still probably strong, and this required an improving brain also. However, tool building was almost certainly taught by example, as it had been for millions of years, and not by verbal instruction. Even today we best learn to use, make, and design tools by example and experimentation and not by verbal instruction. The main advantage of language was that it made possible a flexible, complex organization of human beings which could be passed on from generation to generation. From this time on the direction of evolution was based more and more on competition between groups and not individuals.
Organization and EvolutionNatural selection operated by assuring survival to the most effective hunting group and not the most effective individual, no matter how great his individual powers. Those groups which had the most superior individuals would have been most outstanding, other things being equal. However, the cooperation, sympathy, and discipline of the individual was just as important as his/her technical ability. Those groups which developed the most effective rules of social organization might out-perform another group in which the individual ability was greater. Those groups which became burdened with irrational rules of behavior and superstitions might be at a disadvantage relative to the other less entropic groups.
It was necessary for the group to be mutually supportive and protective, but it could not be burdened for long with defective members who could not pull their own weight or who in other ways increased the group's entropy. Up to the present time the problem of what to do with parasitical human beings is a major ethical question. The Eskimos of a few hundred years ago solved it through group pressure, which drove the old and the crippled to commit suicide by freezing when they were a burden to the group.
In the past, dependent individuals simply died as soon as their parents stopped caring for them. Once group hunting started, there was survival value in nurturing wounded or sick adults who had valuable skills, because their abilities would in time reassert themselves. Even a permanently disabled hunter could be of value to the group if he could keep up with the group, which was constantly on the move, as are all hunters, and at the same time he could make tools and perform other services normally done by non-hunters. Thus, most hunting groups developed a social ethic which increased individual security by assuring group protection to those who became disabled, so long as they could perform valuable services.
A fossil has been found of a Neanderthal man who had lost an arm many years before he died, as indicated by the healed bone. He could not hunt or survive as an individual, but the group protected him for many years because it cherished him. A bond had been formed between him and the group which was similar in its effect to the sexual bond existing between mates. Some anthropologists call this the "hunting bond" [543]. It occurs between men who hunt together and become mutually interdependent. We see its manifestation today in the close friendships that develop between men who fight together as a single unit against a common enemy in wartime. This type of bonding does not seem to ever encompass more than about ten individuals at a time and sometimes many fewer. This is about the same upper limit as the number of adult male hunters that the primitive hunting groups could support. It is the same upper limit that most military organizations in the world today put on the most basic military unit - the squad or patrol. It is the same upper limit on the size of a team in most competitive team sports.
So long as humans lived primarily as hunters, they could not organize into bands totaling much more than 50 individuals. The average band was probably no larger than 32 with about eight adult male hunters. When game became scarce, the bands had to break down into even smaller subgroups possibly even to the individual family unit. This situation caused a change in male-female relations, since the basic polygamous pattern of the hominids for the past four million years - and of many primates in general and our closest relatives, the pongids, in particular - was no longer economically feasible. There was too much survival value in group hunting to have it succumb to the basic polygamous sexual instincts. It was advantageous to have as many male hunters banding together as the game density would support, but there was not enough food for all to have polygamous families. Therefore, man became by necessity monogamous, as are almost all group hunting species. The most dominant males might on occasion have as many as two or three mates, but this was probably rare. The basic structure of hominid society from about 1.5 million years ago to about 10,000 years ago probably consisted mainly in groups of monogamous families, rarely exceeding eight families and having exogamous mating. The Eskimos who survived as group hunters in relative isolation until about 100 years ago had this type of social structure.
Monogamy and Sexual SelectionSexual selection plays a very important part in evolution. As was indicated earlier, the development of sexual reproduction represented a considerable advance in evolution since it increased the variability of the population undergoing natural selection and, in effect, enabled the species to have a much higher mutation rate without significantly increasing the rate of deleterious mutations. So long as the hominids were pursuing the traditional, generalized primate mode of polygamous mating, with the most dominant male having the maximum access to the females, there was no particular evolutionary advantage for the male to be highly selective in choosing one female over another. He simply chose every female that was available, since under the proper circumstances he could literally keep hundreds of females constantly pregnant - although in practice he probably rarely had a harem that exceeded four fertile females, the maximum number of mouths a proficient solitary hunter could support. However, there was considerable evolutionary advantage for the female to choose the most superior male she could find to give her the most superior young and provide the best possible security for her and her offspring, since she could mate with only one partner at a time. Therefore, evolutionary pressure would produce emotional mutations so that the females were more sexually selective than the males. They would show overwhelming preference for the most superior males available to them, as do all primates today. They would also probably show jealously toward rival females who might endanger their security by diluting the protective capacity of the male with too many mouths to feed. Natural selection would favor females who gave overwhelming priority to caring for their own offspring. Therefore, there was already an evolutionary trend in developing monogamy among the female hominids, almost from the beginning of hominid evolution when the females became increasingly dependent on the males. Monogamy was evolutionarily optimal for the female. That is to say, the "fitness" of the female is maximized by monogamy, while the "fitness" of the male is maximized by polygamy. "Fitness" refers to the potential for producing progeny.
Since primate males tend to exercise absolute dominance over the females, the males imposed polygamy on the females. A compromise which was optimal for the species was probably reached, one that emotionally preprogrammed the females to become increasingly intolerant of new rivals as the male built up his harem. Eventually the older females would band together to drive away any new rival whenever the protecting male was absent. As time went on, through natural selection, this intolerance toward new females increased so that the harems became constantly smaller on the average. At the same time, the larger his harem, the less interested the male became in adding to it, so that an evolutionary compromise was reached between the male and female halves of the species which produced an optimal family size from an evolutionary point of view. This was a compromise which would give maximum reproductive advantage to the most superior males while constraining the danger to the females and their offspring from having to compete for food and protection against their immediate rivals. The compromise was one which maximized the expected number of surviving offspring for the male.
As natural selection favored the growing intolerance of females against mating rivals, and the male harem decreased in size, there was an evolutionary advantage for the male to become more selective by choosing as his mates the most superior females, i.e., the strongest, healthiest, and most intelligent females. By the time group hunting became technologically feasible, there was already a strong predisposition toward monogamy among hominids, although it was considerably stronger in the female than the male. The choice of mate had long been a crucial one for the female, and it had become increasingly important for the male. Now it became all-important for both.
The survival of their progeny and their continued evolution depended on the choice of mate for both males and females. The evolutionary trend was toward growing, generalized intelligence. Therefore, sexual selection was probably strongly based on intelligence. Even today the strongest correlate between all measurable characteristics of married couples is their I.Q. [389]. It is not meant to imply by this that I.Q. is synonymous with intelligence, but rather that it is indicative of a limited aspect of intelligence. If a more direct measure of intelligence were available, it would probably be even more highly correlated for married couples. This correlates more strongly than any physical characteristic or even socio-economic background, religion, and values. It is an indication of ancient emotional programming that has been genetically determined for millions of years and is still operating. We see it on another level by the much greater selectivity of women in choosing sexual partners (not necessarily mates) than is shown by men [415, 416]. However, the new epidemics of sexually transmitted diseases are changing this pattern.
The important consequence of monogamous mating through intellectual sexual selection is that it probably increased the evolutionary rate for humanity [389].
Modest polygamy maximizes the evolutionary rate when there is high selectivity on both sides. But emotional preprogramming in the human male is such that he becomes highly indiscriminate when he has the opportunity of unlimited polygamous mating [415]. Only monogamous mating emotionally predisposes him to be highly selective. Therefore, monogamy not only was an essential prerequisite for group hunting, but it probably also accelerated the evolution of the human brain. Monogamy and group hunting together produced a synergistic interaction on the evolution of humanity in general and human intelligence in particular.
Group EvolutionThrough exogamous mating and the development of language, there was a constant flow of information between different hunting bands in the form of genes and machines (tools, language, and social organization). As language developed, hunting bands which were isolated from one another became linguistically differentiated so that it was more difficult to exchange information when they met. This is analogous to the differentiation of isolated species in different environments. In time, there probably developed a hierarchy of communication between persons in which the freest flow of information was within the band, then within the clan, then within the linguistic group, and finally within the race. Taboos probably developed about mating outside the linguistic group. This and geographic separation probably gave rise to racial differentiation among Homo erectus.
As our ancestors continued to fill the earth, they came into conflict with one another concerning territorial rights; there was increasing warfare between bands, clans, tribes, nations, and races in that order. In this context, a "nation" is a set of all persons with a common language. The larger the number of hunters that could mass on one side, the greater the probability of success in battle; but the game supply mitigated against large concentrations of hunters. Therefore, those groups of bands that could develop a type of government for working together against common enemies would be more likely to survive and increase their numbers. This put even more evolutionary pressure on the development of language until the more abstract notion of hierarchical loyalties could be codified so that all persons knew their obligation to their family, their band, their clan, their tribe, and their nation. Probably an instinctive aversion developed, a type of xenophobia, to significantly different races (and as a consequence aversion towhat were viewed as "inferior," i.e., different, cultures), or distinct races probably would not have formed except through absolute geographic isolation. The group hunting Indians of North America had these types of social organizations and behavior patterns, although they had much more evolved machines than group hunters of 200,000 years ago. There is also evidence of mass genocide occurring when very different hominid hunting groups (different in culture or race) came together. In recent times this happened when the Iroquois exterminated the Mohicans, the Polynesians exterminated the Australoid aborigines, the Romans exterminated the Carthageneans, the Zulus exterminated the Bushmen, and, in Lebanon today, the Moslem and Christian Arabs exterminate one another. Similarly Cro-Magnon probably exterminated the European Neanderthals.
The NeanderthalsAbout 400,000 years ago, the hominids began to mutate into two distinct strains, each one of which was more like Homo sapiens than Homo erectus. One, the most generalized, was to lead directly to modern humans; the other led to a widespread and temporarily more successful group, the Neanderthals. The Neanderthals represent the last significant divergence from the main trunk of hominid evolution. Both the sapiens and the Neanderthal strains were to continue developing their brain, but the Neanderthal brain developed along a very different direction [152, 225] as shown here.
Both the sapiens and the Neanderthal strains developed along the direction of becoming more effective hunters. However, the Neanderthals emphasized an increased development in the sensory and motor centers primarily in the rear half of the brain, while the sapiens emphasized an increase in the higher centers - i.e., the speech, imagination, and, above all, ethics centers - in the front half of the brain, particularly the frontal lobes which were twice the size of the Neanderthals' [225]. Homo sapiens' Brain versus Neanderthal's
Both developments were worthwhile. But that of the Neanderthals was more specialized, and it was of the same nature as having more acute vision and stronger muscles. The brain was reaching its limit of maximum size, because of the obstetric problem mentioned earlier, and it could go in two directions. Both changes would increase hunting effectiveness and even tool-making capability. The Neanderthals would become superb athletes with excellent memory for spatial relationships and the ability to discriminate between complex auditory and visual stimuli. They probably had greater manual dexterity than any hominid group before or since, and were capable of making excellent tools, considerably superior to those of Homo erectus. However, they probably had very limited linguistic ability. A recent analysis of the anatomical structure of the mouth cavity and larynx of Neanderthal man indicates that he could probably not even physically form the complex sounds of modern language. Perhaps even more disastrous at this stage of evolution was the Neanderthal's much less developed frontal lobes, which are the cortical centers of forethought, imagination, invention, and ethics. These are essential to create human organization. His superior sensory and motor equipment probably gave Homo neanderthalensis a temporary advantage over Homo sapiens in hunting ability, but the basic pattern of human evolution was not in this direction; it was in the direction of increasing ability to use, build, and invent machines. Invention is the most generalized ability that any species can have. It was an ability in which Neanderthal was relatively deficient, because of his brain structure.
Still, for several hundred thousand years Neanderthal man was able to expand over much of the earth and to completely dominate Europe. In Europe, which became totally glaciated during this period, Neanderthal man survived until about 30,000 years ago. Once he began specializing his brain in the direction of sensory acuity and motor dexterity, the rest of the body followed and he developed into a squat, very heavily-muscled creature with massive, bony eyeridges for protecting his eyes. The healthy, classic Neanderthal male was about 5 foot 6 inches and weighed about 200 pounds. (In comparing hominid subspecies and races it is convenient to consider the males instead of females because the males are more highly differentiated than the females.) Still, he developed very elaborate tools and great hunting skills so that he could kill the enormous cave bears of Europe; these he massacred in great numbers, possibly for food and to take over their caves, but partially it seems for religious purposes [152, 353].
Neanderthal man buried his dead in complex rituals which included decorating the body and providing it with tools. He almost certainly possessed a rudimentary art. He also seems to have practiced ritualistic cannibalism, which may have been limited to eating only the corpses of honored members of the hunting band or family who had died of natural causes. (He was probably a gentle creature who did not prey on his own kind, as is evidenced by his caring for maimed persons.) The practice currently occurs among certain Papuan tribes in New Guinea, which concentrate on eating the brains of honored ancestors. Neanderthals also seemed to perform trepanning rituals on each other in which the living brain was exposed, probably to let out evil spirits or let in good ones. Some Neanderthals survived this ceremony, as evidenced by the healed bone in the fossil skulls.
All in all, Neanderthals were quite human, particularly in their emotional makeup and solicitude for injured members of the band. They were religious, artistic, industrious, compassionate, and courageous. Their specialized brain was 25% larger than modern man's. However, they could not have been as inventive, well-organized, or ruthless as our ancestors who began as a small, invading minority from North Africa among the European Neanderthals and who in only a few thousand years had completely replaced them in what may be the first case of mass genocide on earth.
Cro-MagnonCro-Magnon is not only fully human, i.e., a Homo sapiens, but he seems on the average to have been superior to modern man in brain and body. The typical Cro-Magnon male was a superb physical specimen averaging close to six feet in height and weighing about 200 pounds. More importantly, he had a fully modern brain almost 20% larger on the average than a typical modern human brain! The implications of and reasons for this apparent degeneration in Homo sapiens will be discussed later.
The earliest Homo sapiens-like fossil - Heidelberg man - dates from 400,000 years ago and was found in Central Europe. There may have been an island land bridge formed across the Mediterranean by way of Malta during this period as the ocean's waters were taken up in the glaciers that were to cover Europe. By 100,000 years ago the Neanderthals were distributed all over Eurasia and Africa. It seems that the Neanderthal adaptations were primarily those of a hunter to extreme cold. He may have been very hairy, but most likely he made clothes out of skins for protection. The late European and Asiatic Neanderthal probably had a way of life very similar to the Polar Eskimos of a few hundred years ago, although they lacked many important Eskimo tools, particularly snowshoes and needles, modern language, and complex social organization. Cro-Magnon, on the other hand, seems to have evolved in the much more temperate climate and conditions of Africa, which enabled him to remain more generalized and then invade Europe en masse about 45,000 years ago. Recent genetic evidence indicates that all humans are descendants of a single Homo sapiens Sub-Saharan African woman who lived about 200,000 years ago. All other hominids were apparently exterminated by this invading African strain of Homo sapiens [901].
Forty-five thousand years ago, Europe was completely dominated by the Neanderthal. Within 15,000 years all Neanderthals had disappeared. We know that there was very little interbreeding between the Neanderthals and the Cro-Magnon in Europe because there are no intermediate fossils. This might at first seem somewhat strange because, although it is conceivable that the Cro-Magnon men might have killed the Neanderthal males in dispute over hunting territory, it is characteristic of primitive men to subjugate the women of the conquered peoples and breed with them. If this had happened, then classical Neanderthal genes would have entered into the mainstream of human evolution through Neanderthal women and we would find fossils of intermediate hybrids. But this is not the case. Therefore, this was a true case of genocide. The Neanderthal women were either killed or left to starve. This might have occurred because of the evolutionary pressure for monogamous mating and the intolerance of Cro-Magnon women, but most likely it was due to the fact that the Cro-Magnon men were not sexually attracted to Neanderthal women. Knowing the male Homo sapiens' proclivity toward polygamy, or at least eclectic sexual adventure [415], this implies that the European Neanderthals had mutated into a subspecies that was almost universally repulsive to our ancestors. The Neanderthal men themselves might not have been so discriminating, but they were in no position to choose, and became extinct.
In the more temperate parts of the world, however, intermediate fossils between Neanderthal and Homo sapiens have been found from this period. It seems that there was strong evolutionary pressure in both the Neanderthal and the Cro-Magnon direction 200,000 years ago. Occasionally bands which had not gone too far in the sapiens or the Neanderthal direction would come together, interbreed, and produce hybrids. Through them we may all carry some Neanderthal genes, although the Cro-Magnon strain clearly predominates. The recent genetic finding that all humans are descendants of a single sapiens female that lived in Africa about 200,000 years ago is further evidence that the Neanderthals were exterminated when the African strain of Homo sapiens began to invade Eurasia over 100,000 years ago.
The European Neanderthal, having been isolated by glaciation from the rest of humanity for 30,000 years or more, carried the Neanderthal specialization to its limit. The Cro-Magnon evolutionary characteristics were probably developed in relative isolation from Neanderthal types for 100,000 years or more, and when the two subspecies met they no longer had enough in common to interbreed. It is even conceivable that they were not mutually fertile, although this seems more far-fetched, since even lions and tigers are mutually fertile, as are the most divergent subspecies of dogs. The differences between the most divergent human races of today are not enough to prevent interbreeding. But the differences between the European Neanderthal and Cro-Magnon man were far greater than those between any two existing races, so there is a slight chance that they could not interbreed, in which case our ancestors of this period might not necessarily be guilty of deliberate genocide.
However it was that Cro-Magnon contributed to the extermination of Neanderthal, by 35,000 years ago he had a flourishing group hunting culture all over Europe. He had elaborate religious customs and was highly creative. He developed increasingly more complex tools, including the bow and arrow, created great works of art, and laid the foundations for the modern languages. Neanderthal man might have copied but did not invent the bow and arrow and other Cro-Magnon tools; if he had invented this superb weapon, it is he who would have exterminated Cro-Magnon and not the other way around. We further note that the Australian aborigines never invented the bow and arrow, although they did have the spear thrower.
The Basques may be the closest genetic and linguistic descendants of the Cro-Magnon. They (1) are anatomically similar to Cro-Magnon, (2) are the only group of persons to have over 60% Rh-negative blood types (an ancient characteristic), (3) live in the same isolated regions about the Pyrenees as the early Cro-Magnon, and (4) have a unique language unrelated to all other languages. However, the Cro-Magnon of yesterday, like the Basques of today, have interbred with the succeeding waves of humanity who came to Europe from Asia and Africa to produce all modern European types. The Europeans are only one of the many races which constitute Homo sapiens.
Homo sapiensAll living hominids are members of the same species, Homo sapiens. The earliest clearly Homo sapiens fossil is about 75,000 years old. Some anthropologists classify Homo neanderthalensis as Homo sapiens. However, for the reasons discussed in the previous two sections, this does not seem to be proper, particularly since there was so little interbreeding, for whatever reasons, between Homo neanderthalensis and Cro-Magnon (who existed in the same areas for over 15,000 years).
The evolution of Homo sapiens has been almost entirely cultural, not biological. Indeed, we seem to be degenerating biologically. Homo sapiens has carried the main trend of hominid evolution, i.e., the increasing ability to create and use machines, to its logical conclusion and has become completely dependent on machines; this is one cause for the degeneration of the body. A certain amount of biological adaptation has occurred, as shown in the diversity represented by the five basic racial types, namely the Mongoloid, Negroid, Caucasoid, Australoid, and Capoid.
The mongoloid characteristics are a biological adaptation to extreme cold (epicanthean eyefolds, coarse insulating hair, abundance of subcutaneous fat in the face, low surface area to mass ratio, relatively few sweat glands, etc.). The negroid adaptation is one to extreme heat (dark skin, relative abundance of sweat glands, ventilated sun helmet of kinky hair, high surface area to mass ratio, etc.). The nilotic negro is a desert dweller and the pygmy a forest dweller. Both seem to be descendants of a common intermediate type. The caucasoid characteristics are not so much an adaptation, with the possible exception of the long, thin, air-warming nose, as a degeneration - although the pale skin is a degeneration which gives advantage in a cool, cloudy environment by increasing the amount of vitamin D synthesized in the body by the action of sunlight. The Australian Aborigine is the Homo sapiens most like our ancestors of 100,000 years ago, although he has a much smaller brain and thicker skull than Cro-Magnon man or any other living race of man, and indeed has many characteristics of Homo erectus [30]. Racial differentiation between negroids and other human populations began about 115,000 years ago (-- 34,000 years) and about 40,000 years ago (-- 15,000 years) between caucasoids and mongoloid types [901]. There has been no absolute genetic isolation between races in the last 100,000 years except for the Australian Aborigines, who were genetically isolated for at least 30,000 years.
Many other human types represent more generalized races which have not specialized as much as those noted above or which are hybrids of several more specialized races. In terms of their biological success, the caucasoids are first (55% of the hominid biomass); then come the mongoloids (37% of the hominid biomass); together the mongoloids, caucasoids, and their various hybrids account for over 92% of the human species. The negroids account for less than 7% of the human species, and the Australoids and capoids are less than half of one percent of today's humanity [30, 172]. It should be pointed out that the large increase in caucasoids is a phenomenon of the last 300 years. Up to that time, the mongoloids had been in the ascendancy, competing off and on for the last 10,000 years with the caucasoids for domination of the hominid biomass.
Since the advent of Homo sapiens, evolutionary competition has become increasingly bloody and direct between the various human subgroups. The basis of this competition has been the ability to create and use the complex machines which are the means of modern hominid evolution. The machines are used to control the natural resources - hunting grounds, arable lands, minerals, people, etc. The most important machine for the last 10,000 years or possibly longer has been human organization. For when humans could organize themselves into a progressive society which enhanced the basic evolutionary pattern of increasing the coherent information within the biomass, then they could create ever greater machines and increase their numbers. There was a constant interplay and synergistic interaction between the machines of language, organization, and tools. For humans, evolution is now almost entirely psychosocial.
Psychosocial EvolutionPsychosocial evolution is unique to the hominids. There is no evidence that any other animal group evolves or has ever evolved this way. As was pointed out earlier, the basic process of evolution involves an increase in the amount of true information in a system. In general, random mutations decrease the true information in the system and increase entropy, just as substituting words at random into a coherent book decreases the true information in it. However, if (1) books could reproduce much faster than they were being disrupted by random changes, and (2) those books which had the most coherent information reproduced much faster than those that had least information, and (3) the most incoherent books did not reproduce at all, then we might have a library of evolving books. However, this would normally seem like a very inefficient way of improving our library, even if we had books which followed the rules of natural selection. A much better way is to make nonrandom, directed changes in the books. This is precisely the process of psychosocial evolution.
We can look at a species as a library of true information. For all species, except Homo sapiens, this information is contained entirely in the collective DNA of the species, which clearly contains much redundant information, although each individual is unique. This DNA is constantly mutating and recombining into new patterns. Through natural selection, any species can continue to evolve until it becomes overly specialized. Then it becomes extinct. This has been the pattern of all evolution, including the hominids'. By building machines, humans increase their generalized intelligence and avoid specialization.
Early in hominid evolution our ancestors began to store extragenetic information. This was the information on how to build and use machines. This information was passed from one generation to the next; but it had to be learned after birth. No hominid has ever been born knowing how to build any machine, although they could have been born with a genetically determined body structure, e.g., a better brain, which made it easier for them than others to learn how to use and or invent machines. Invention was the essential psychosocial mutation which made this new type of evolution possible.
An invention is a new machine which can increase the "fitness" of the species. "Fitness," we recall, refers to the potential of a species or of an individual for producing progeny. Fitness is a direct function of nutrient-gathering, mate-finding, and reproducing capability. An invention is produced by rearrangement of the physical, biological and/or psychosocial environment into a new pattern which produces a new and/or more effective machine. For example, tools involve physical invention, medical care involves biological invention, language and social structure involve psychosocial invention. So long as the hominids were organized into small groups, the benefits of invention were limited almost entirely to the family in which the inventors lived and their descendants. Therefore, the ability to invent and pass on the knowledge of this invention to one's progeny was the major direction of the evolutionary change, once full upright posture was achieved.
The critical inventions which are responsible for human evolution are many and interdependent, e.g., tools, language, government, agriculture, science, mathematics, computers, radios, etc. Machines evolve from generation to generation analogously to a species. Eventually primitive technologies disappear and are replaced with more modern technologies which could not have existed without the previous now-lost technologies (e.g., very few living humans can make a fire without matches or other modern machines). Technology is the process for designing, building, and using machines. Therefore, the more evolved the hominids became, the more dependent they were on their nongenetically inherited technology - until today the typical human can no longer exist without either machines or accumulated knowledge.
The machines themselves became extensions and amplifiers of the body and serve the same functions as the more specialized organs in other species. A club substitutes for the massive arms of the pongids; a sharp stone substitutes for their sharp, large canines. A spear substitutes for the horns of the grazing animal; a knife, for the claws and fangs of a carnivore. A bow and arrow has no precise analog in nature, but it is superior under many circumstances to any combination of horn, fang, and claw. It is an extension of our stone-throwing ability. Today we build machines that enable us to fly faster and farther than any bird, telescopes and microscopes which enable us to see better than any unaided eye, electronic chemical sensors which enable us to smell better than any bloodhound, and language which enables us to process, store, and exchange information in a way which is analogous to the nervous system of our body. This capability is further amplified by the invention of computers and electronic communicators (radio and television). The pattern which emerges is that humanity is forming an interdependent collective entity with analogous but greatly amplified characteristics of its own body. Humanity is a unified whole of persons, machines, and knowledge. Each part is essential to the survival of the other. Together all the parts create a collective entity which can transcend biological limitations.
Biological LimitationsThere are evolutionary limitations inherent to the structure of life as we know it. The major limitation is in the amount of DNA a cell can contain. The DNA containable in a cell is limited because the overall size of a cell is limited by the surface area to mass ratio and other physicochemical relationships. The surface area to mass ratio must be at least equal to the critical value for the human ovum, or the cell will not be able to exchange gases with its environment at a rate adequate to maintain a viable metabolism. Therefore, a cell cannot be more massive than about 10-6 gms, and since there must always be less DNA in a cell than its total mass, the complexity of life is limited to the amount of information that can be stored in 10-6, or fewer, gms of DNA. Actually this is a very crude upper limit to the complexity of life, since (1) the actual amount of DNA in a cell must double before reproduction, (2) much of the cellular information must be redundant and/or used for self repair, or the cell will be highly unstable, and (3) the DNA is less than 10 percent of the mass of any cell. Therefore, a more realistic upper limit on the maximum amount of nonredundant DNA available for structuring the complexity of any given individual is probably closer to 10 gms. Whatever the case, it is clearly a finite amount of a fairly small order.
There are also other limitations such as the practical limitations on the size of the nervous system and the amount of redundancy that must be put in the nervous system to ensure stability and reliability. However, just taking the mass of nonredundant DNA as the limit to the genetic information that life can contain, we see that evolution solely through genetic mutations is quite limited, and probably not much more than the complexity of a human being is obtainable through this means. Humanity is already a highly unstable species on the verge of suicide [280]. However, the amount of coherent information available to humanity through psychosocial evolution is virtually unlimited. Humanity has continued and can continue to evolve culturally, although our bodies on the average seem to be decaying when compared with Cro-Magnon. The critical words are on the average.
The largest hominid cranial capacities ever observed occur in modern humans, not Cro-Magnon. Therefore, although the average human brain may be decreasing in size - and recall that there is more to complexity than size - the maximum complexity of the human brain probably is as great as or maybe even greater than it has ever been. The reason for the decreasing average brain size, while the maximum brain size is at least as large as it has ever been, is that as humanity invented agriculture (probably a female invention, as shown in Chapter 4) and civilized society, it became possible for relatively defective human beings to survive and reproduce by performing simple specialized services for the society as a whole. These persons could not have survived in a competitive hunting society where they would have to perform many different functions - tool maker, hunter, warrior, cook, tailor, physician, etc. - but they could survive as a specialized part of a much more complex society. As society becomes more complex, it becomes possible for ever more defective individuals to survive and reproduce, until survival and reproduction are possible today for large numbers of total parasites who are incapable of even caring for themselves and would quickly die if someone less defective did not constantly provide for them and their children.
At the same time, as machines take over more and more of the specialized functions of human beings, it becomes possible for ever fewer persons to invent, produce, use, and maintain all the essential machines of society. Therefore, the range of human capabilities becomes broader while the average becomes lower, and increasing numbers of human beings begin to live parasitical existences in an increasingly automated society. It is possible for human evolution to continue, at least up to a point, at an increasing rate, even though the vast majority of the human race is degenerating.
Up to now the transcendence of the biological limitations on human evolution through psychosocial evolution has enabled humanity to evolve at an ever increasing rate and to compensate for the increasing entropy in pure biology. However, this is not a simple linear process, and increasing biological entropy can interact with all other components of human information to produce psychosocial entropy which can eventually reverse human evolution and lead to extinction, just as it has for so many other species. In order to see that this is the case, we consider the evolution of mind.
![]()
© John David Garcia, 1991, All rights Reserved.